On October 21st, 2021, Prof Johan Storm gave a presentation about the history, progress, and problems in Consciousness science for the Norwegian Academy of Science. The presentation available (in Norwegian) here: BEVISSTHETSFORSKNING i Det Norske Videnskaps-Akademi
All posts by Bjørn Erik Juel
THE DREAMING BRAIN On Sleep, Dreaming and Consciousness
Forum for Consciousness Research and The Norwegian Academy of Science and Letters invite you to an open meeting on the science and philosophy of sleep, dreaming and consciousness.

Schedule:
11.00 – 11.05: Introduction by Johan F. Storm, Neurophysiology, University of Oslo
11.05 – 12.05: Antti Revonsuo: On the fundamental nature of dreaming: From dream philosophy to consciousness science.
12.05 – 12.20: Coffee break
12.20 – 13.20: Francesca Siclari: Conscious experience in sleep: a high-density EEG assessment
13.20 – 13.50: Discussion and questions from the audience
Dr. Antti Revonsuo (Professor of Cognitive Neuroscience, University of Skövde) is a Finnish cognitive neuroscientist and philosopher of mind. He seeks to understand consciousness as a biological phenomenon and his work focuses on altered states of consciousness with dreaming in particular. He is well known for his Threat Simulation Theory, which states that dreams serve the biological function of rehearsing possibly threatening situations in order to aid survival.
Dr. Francesca Siclari (Lausanne University Hospital) is a medical neuroscientist specialized on consciousness and dreams. She seeks to understand consciousness based on dream content and their neural representation in the human brain. Her work together with Giulio Tononi is best known for describing the first neural correlates of dream experiences in the source reconstructed EEG.

The year of consciousness research: Washington, New York, Barcelona, and Krakow
In the year past, there have been several interesting events related to consciousness research.
Washington – Society for Neuroscience
A symposium titled «Neural Correlates of Consciousness: Progress and Problems» was organized at the annual conference for the Society for Neuroscience (SfN). Melanie Boly, Marcello Massimini, Cyriel Pennartz, and Melanie Wilke presented different experimental approaches and theoretical frameworks relevant for consciousness research, and the themes of the symposium were presented in a conference paper in the Journal of Neuroscience. As this was the first major symposium covering consciousness research specifically in the 47 year histort of the Society for Neuroscience, it can bee seen as a milestone for the maturity of the field. The symposium was chaired by Johan Storm, and was supported by the Human Brain Project.
New York – Animal Consciousness Conference
The Center for Mind, Brain and Consciousness (in conjunction with the Center for Bioethics and Animal Studies) at New York University hosted a conference on Animal consciousness, with series of distinguished speakers presenting and discussing the latest results and ideas from this field, including ethology, neuroscience, and philosophy of mind. More information about the conference, along with program, abstracts, attendance, and videos of talks can be found at the following website.
Barcelona – HBP Understanding Consciousness, a Quest for the 21st Century.
Consciousness research entered sunny Barcelona in late June. In two packed days there was talks by eminent neuroscience researchers and philosophers such as David Chalmers, Ned Block, Karl Friston, Jean-Pierre Changeux, Wolf Singer, Emery Brown, Olaf Blanke, Marcello Massimini, Rodolfo Llinas, Larissa Albantakis, Nico Schiff, Melanie Wilke, and many other excellent speakers. For an overview of speakers, talks, abstracts, and more, go to the website.
This was the first in a series of large Human Brain Project conferences, and was organized by Johan Storm together with other HPB members.
Krakow – Association for the Scientific Study of Consciousness
A short week after the Human Brain Project conference on consciousness in Barcelona, the ASSC opened the doors in beautiful Krakow. For almost a week, attendees got their fair share of talks, posters, workshops, and symposiums, on various topics within the interdisciplinary field of consciousness research. Summarizing everything that happened in a few sentences wouldn’t do it justice, so make sure you check out the web site or their Facebook page.
We hope to publish more information about these events and others like them in the time to come.

Large HBP conference on CONSCIOUSNESS in Barcelona, June 21-22, 2018
UNDERSTANDING CONSCIOUSNESS – a scientific quest for the 21st Century.
A 2-day scientific conference linking the Human Brain Project and external neuroscience communities.
Auditorium in CaixaForum, Barcelona, Spain
June 21 – 22, 2018
First International HBP Conference (see Human Brain Project website)
How does our subjective experience emerge from the brain? How does consciousness relate to the physical world? These age-old, deep questions are now at last being addressed directly and broadly by neuroscience and will become crucial in the decades to come.
The spectrum of consciousness-related conundrums is rapidly expanding: we are saving islands of human brain from devastating injuries, growing cerebral organoids in a vat, and building intelligent machines that perform faster and better than any healthy subject, just to mention a few.
Where does consciousness arise? Where is the boundary between insentient matter and a spark of subjectivity? Society needs to be scientifically and culturally prepared to face these emerging questions.
To do so, an approach with the broadest scope is needed: diverse theoretical frameworks, brain anatomy, physiology, and chemistry across scales and species, detailed and large-scale computer simulations, deep learning, neuromorphic computing, robotics, clinical neurology, anaesthesiology, psychology, behavioural, computational, and philosophical analysis must interact and blend on a single infrastructure.
These ingredients are naturally present in the Human Brain Project, and this workshop, stirred by world-leading scholars in the field, is where they come together for the first time.
Registration details will be available in the New Year.
Draft program and preliminary list of external speakers (further speakers and final program to be confirmed)
June 21, AM:
(1) Fundamental aspects, including theories of consciousness (Day1; 9-13):
– Larissa Albantakis: Dept of Psychiatry, University of Wisconsin-Madison, USA.
– Ned Block: Department of Philosophy, New York University, USA.
– David Chalmers: Department of Philosophy, New York University, USA + Centre for Consciousness, Australian National University, Australia.
June 21, PM:
(2) Neurobiological mechanisms and correlates of consciousness (Day1; 14 -18):
– Catherine Tallon-Baudry: Cognitive Neuroscience Laboratory, Institut National de la Santé et de la Recherche Médicale (INSERM)-École Normale Supérieure (ENS), Paris, France.
– Wolf Singer: Department of Neurophysiology, Max Planck Institute for Brain Research Frankfurt am Main, Germany ; Ernst Strüngmann Institute for Neuroscience in Cooperation with Max Planck Society Frankfurt am Main, Germany ; Frankfurt Institute for Advanced Studies, Johann Wolfgang Goethe University Frankfurt am Main, Germany.
– Nao Tsuchiya: School of Psychological Sciences, Monash University, Clayton, Australia + Monash Institute of Cognitive and Clinical Neuroscience, Monash University, Clayton, Australia.
– Rodolfo Llinas: Neuroscience Institute, Departments of Physiology, Neurology, and Psychiatry, and Center for Neural Science, New York University, New York, USA.
June 22, AM:
(3) Models, simulations, and emulation of consciousness (Day2; 9-13):
– Sean Hill: Neuroinformatics division, Campus Biotech, Geneva, Switzerland + Laboratory for the Neural Basis of Brain States, École Polytechnique Fédérale de Lausanne, Switzerland.
– Karl Friston: Wellcome Trust Centre for Neuroimaging, Institute of Neurology, University College London, London, UK.
– Fabrice Wendling: INSERM, U642, Université Rennes 1, Rennes, France.
June 22, PM
(4) Clinical, ethical, and societal implications of consciousness research (Day2; 14 -18):
– Nicholas Schiff: Feil Family Brain and Mind Research Institute + Dept of Neurology, Weill Cornell Medical College, New York, USA.
– Emery Brown: Department of Anesthesia, Critical Care and Pain Medicine, Massachusetts General Hospital, Harvard Medical School, Boston, MA, USA + Department of Brain and Cognitive Science, Massachusetts Institute of Technology, Cambridge, MA, USA.
– Melanie Wilke: Department of Cognitive Neurology, University Medicine Goettingen, Goettingen + German Primate Center, Leibniz Institute for Primate Research + German Research Foundation (DFG) Center for Nanoscale Microscopy and Molecular Physiology of the Brain (CNMPB), Georg-August-Universitaet Goettingen, Goettingen, Germany.
– Olaf Blanke: Laboratory of Cognitive Neuroscience, Brain-Mind Institute, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland + Center for Neuroprosthetics, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland.
Organizing program committee
• Alain Destexhe (SP4, Paris)
• Kathinka Evers (SP12, Uppsala)
• Marcello Massimini (SP3, Milan)
• Olivia Gosseries (SP3, Liege)
• Cyriel Pennartz (SP3, Amsterdam)
• Johan Storm (SP3, Oslo; coordinator)
Advisor group
• Jean-Pierre Changeux (SP12, Paris)
• Christof Koch (President and Chief Scientific Officer of the Allen Institute for Brain Science, Seattle, USA)
Public ROBOT presentations by Tony Prescott/Martin Pearson

Symposium on Consciousness at the Society for Neuroscience in Washington DC, 15 Nov. 2017
Neural Correlates of Consciousness:
Progress and Problems
https://www.sfn.org/annual-meeting/neuroscience-2017

Theme H: Cognition
Neural Correlates of Consciousness: Progress and Problems – Johan Storm
Chair: Johan Storm, PhD
University of Oslo
Co-Chair: Melanie Boly, MD, PhD
University of Wisconsin-Madison
University of Oslo
Co-Chair: Melanie Boly, MD, PhD
University of Wisconsin-Madison
Date & Time: Wednesday, November 15, 2017 1:30pm – 4pm
Location: Ballroom B
CME: 2.5
Consciousness research is developing rapidly. Using evidence from brain injury in patients and physiological and behavioral studies in humans and related animals (single neuron, fMRI, EEG, TMS, intracranial recordings), the symposium will highlight how different conscious states and contents arise in the brain. Speakers will discuss different experimental approaches and theoretical frameworks as well as the medical and ethical relevance of this area.
Abstract:
The nature of consciousness is widely regarded as one of the great challenges in science. Over the past decade there has been substantive empirical and theoretical progress in this field. This symposium presents results from this recent surge in consciousness research.
Melanie Boly will review evidence of frequent dissociation between consciousness and responsiveness in patients with brain damage. She will present recent evidence that after ruling out confounds, the anatomical neural correlates of consciousness are primarily localized to a posterior cortical hot zone, rather than to a fronto-parietal network involved in task performance and report, and discuss the potential clinical applications of these findings.
Marcello Massimimi will describe the rationale and validation of the perturbational complexity index (PCI), a theory-driven empirical metric designed to gauge the brain’s capacity for integrated information. He will show how this index can be employed at the bedside to assess and stratify unresponsive patients, independently of sensory processing and motor/executive functions, and will highlight physiopathological implications.
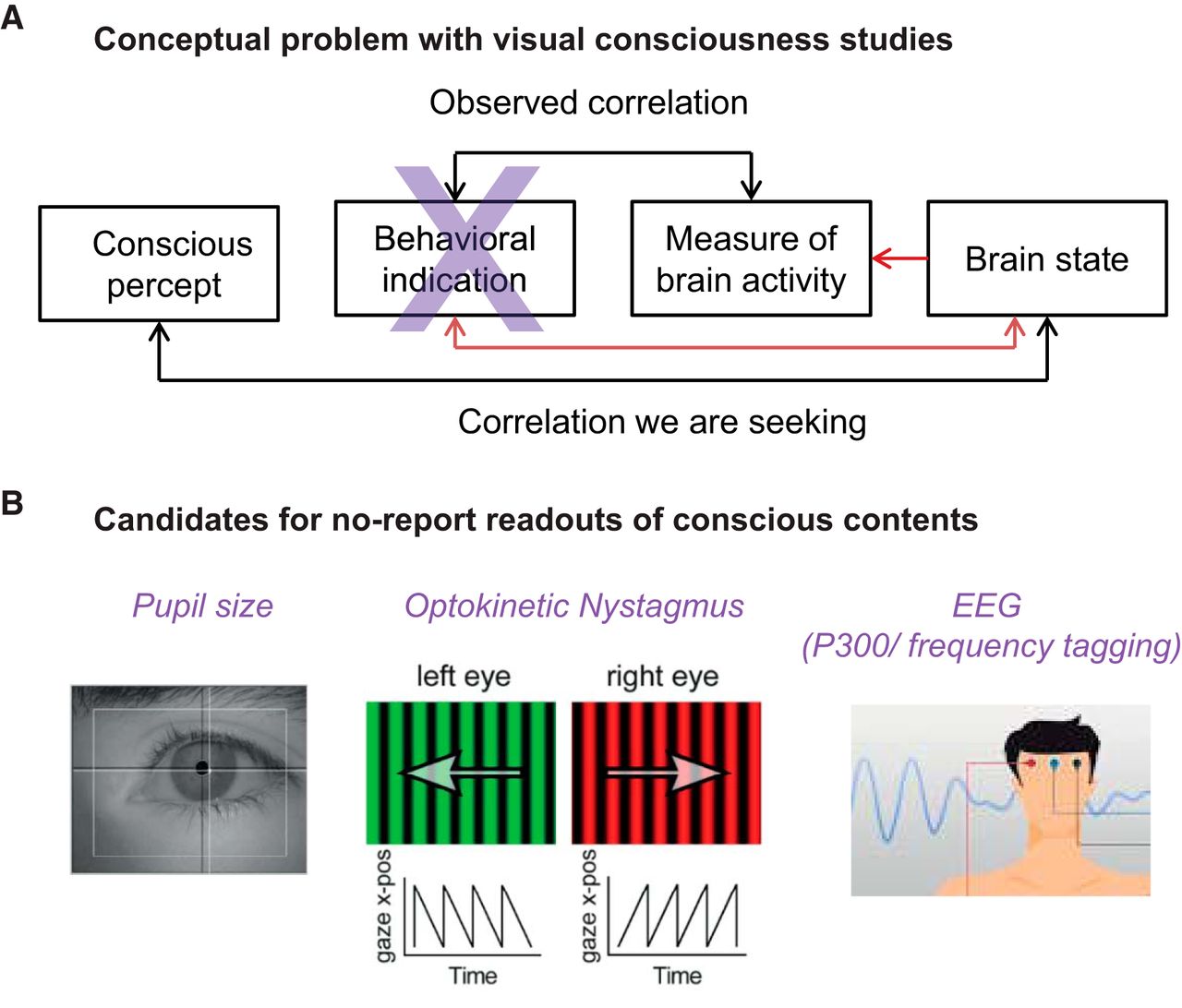
Melanie Wilke will discuss how to disentangle conscious perception from decision making and visuomotor processes. Drawing conclusions from electrophysiological and fMRI experiments in monkeys and humans, she will address which signals and brain regions continue to correlate with conscious perception without the requirement of a behavioral report. She will also show consequences of parietal and thalamic pulvinar perturbations on conscious perception versus visuomotor decisions. Finally, she will discuss how to avoid or control for report-related confounds in future studies of conscious perception.
Cyriel Pennartz will focus on the theoretical delineation of requirements for animal brains capable of sustaining consciousness. Next, he will review recent advances in uncovering neuronal population correlates of visual stimulus detection, which is considered an important component of sensory awareness. Finally, he will zoom out from visual cortex to larger interconnected neural systems, probed with multi-area ensemble recordings to investigate changes in local and long-range functional connectivity across the sleep-wake cycle.
http://www.abstractsonline.com/pp8/#!/4376/session/91



https://www.ncbi.nlm.nih.gov/pubmed/29118218
J Neurosci. 2017 Nov 8;37(45):10882-10893. doi: 10.1523/JNEUROSCI.1838-17.2017.
Consciousness Regained: Disentangling Mechanisms, Brain Systems, and Behavioral Responses.
Abstract
How consciousness (experience) arises from and relates to material brain processes (the “mind-body problem”) has been pondered by thinkers for centuries, and is regarded as among the deepest unsolved problems in science, with wide-ranging theoretical, clinical, and ethical implications. Until the last few decades, this was largely seen as a philosophical topic, but not widely accepted in mainstream neuroscience. Since the 1980s, however, novel methods and theoretical advances have yielded remarkable results, opening up the field for scientific and clinical progress. Since a seminal paper by Crick and Koch (1998) claimed that a science of consciousness should first search for its neural correlates (NCC), a variety of correlates have been suggested, including both content-specific NCCs, determining particular phenomenal components within an experience, and the full NCC, the neural substrates supporting entire conscious experiences. In this review, we present recent progress on theoretical, experimental, and clinical issues. Specifically, we (1) review methodological advances that are important for dissociating conscious experience from related enabling and executive functions, (2) suggest how critically reconsidering the role of the frontal cortex may further delineate NCCs, (3) advocate the need for general, objective, brain-based measures of the capacity for consciousness that are independent of sensory processing and executive functions, and (4) show how animal studies can reveal population and network phenomena of relevance for understanding mechanisms of consciousness.

Review article on Consciousness in The Journal of Neuroscience
A Review article on Consciousness was recently published in The Journal of Neuroscience (8 Nov. 2017).
“Consciousness Regained: Disentangling Mechanisms, Brain Systems, and Behavioral Responses”
Johan F. Storm, Mélanie Boly, Adenauer G. Casali, Marcello Massimini, Umberto Olcese, Cyriel M.A. Pennartz and Melanie Wilke
The Journal of Neuroscience, 8 November 2017, 37(45):10882-10893;doi:10.1523/JNEUROSCI.1838-17.2017

(http://www.jneurosci.org/content/37/45/10882?etoc=)
Abstract:
How consciousness (experience) arises from and relates to material brain processes (the “mind-body problem”) has been pondered by thinkers for centuries, and is regarded as among the deepest unsolved problems in science, with wide-ranging theoretical, clinical, and ethical implications. Until the last few decades, this was largely seen as a philosophical topic, but not widely accepted in mainstream neuroscience. Since the 1980s, however, novel methods and theoretical advances have yielded remarkable results, opening up the field for scientific and clinical progress. Since a seminal paper by Crick and Koch (1998) claimed that a science of consciousness should first search for its neural correlates (NCC), a variety of correlates have been suggested, including both content-specific NCCs, determining particular phenomenal components within an experience, and the full NCC, the neural substrates supporting entire conscious experiences. In this review, we present recent progress on theoretical, experimental, and clinical issues. Specifically, we (1) review methodological advances that are important for dissociating conscious experience from related enabling and executive functions, (2) suggest how critically reconsidering the role of the frontal cortex may further delineate NCCs, (3) advocate the need for general, objective, brain-based measures of the capacity for consciousness that are independent of sensory processing and executive functions, and (4) show how animal studies can reveal population and network phenomena of relevance for understanding mechanisms of consciousness.
Figures:
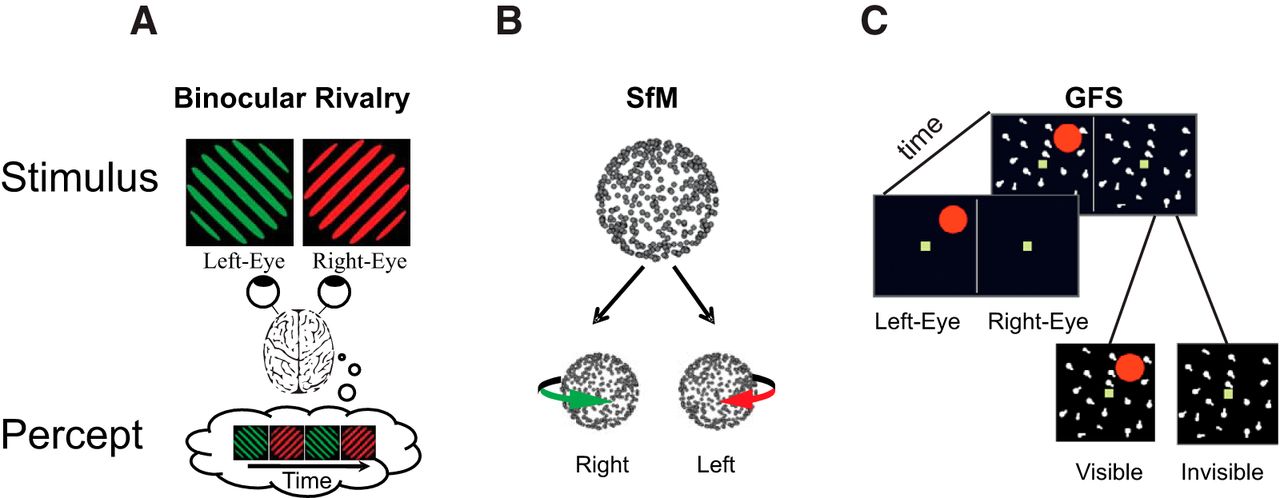
Examples of bistable stimuli. A, In binocular rivalry, two stimuli are shown to different eyes and perception wavers between left and right eye stimuli (Blake and Logothetis, 2002). B, Ambiguous structure-from-motion (SfM) stimulus. Dots moving back and forth on a flat screen, without perspective cues to differentiate between front and rear surfaces, induce the perception of a 3D rotating object that periodically switches direction. (Sterzer et al., 2009) C, Generalized flash suppression. A target stimulus (red dot) is shown parafoveally followed by the onset of a moving surround, causing the red target to disappear in ∼50% of trials (Wilke et al., 2003).
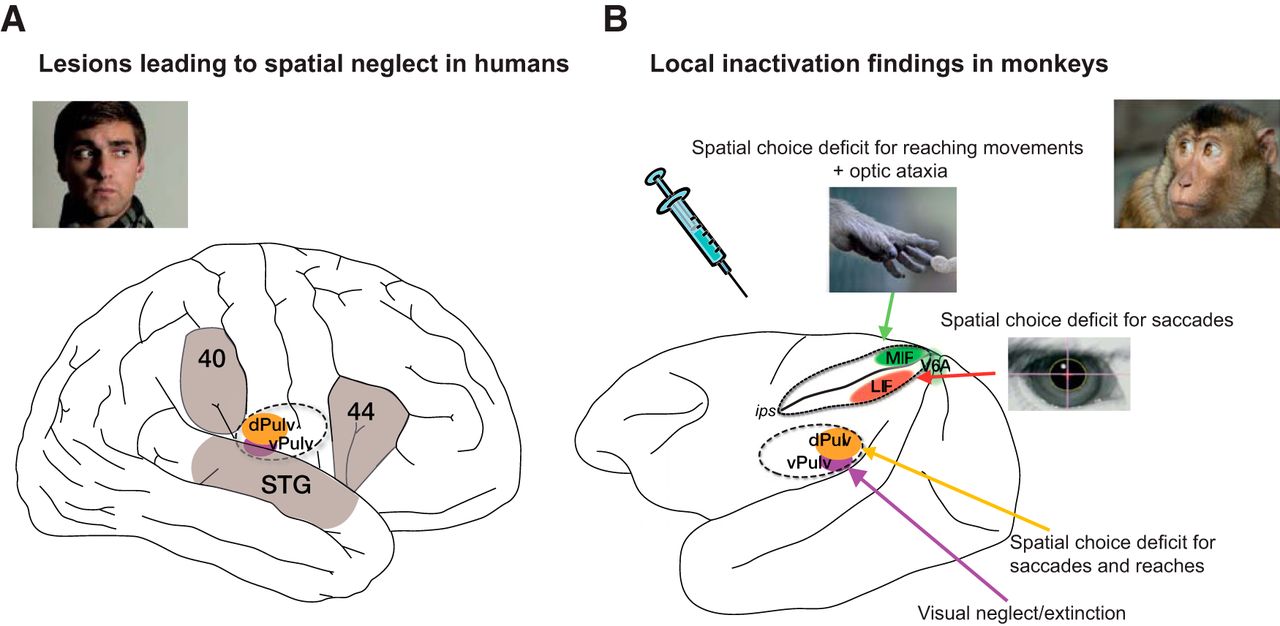
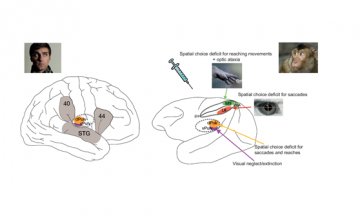
A, Major cortical and subcortical brain regions where lesions lead to spatial neglect in humans (left), and corresponding recent experimental results in monkeys (right) (lateral view). In humans, lesions in frontal (Brodmann area 44), inferior parietal cortex (Brodmann area 40), superior temporal gyrus (STG), basal ganglia, and pulvinar have been reported to lead to spatially biased behavior that might appear as a visual consciousness deficit (Karnath, 2001). B, Recent pharmacological inactivation studies in monkeys have shown primarily effector-specific spatial deficits after lesions in parietal subregions such as the lateral intraparietal area (LIP, red shading) and the parietal reach region (PRR, which includes the medial intraparietal area MIP and area V6A, green shading). Dorsal pulvinar (dPULV, orange shading) inactivation leads to spatial orienting bias for both eye and hand movements which can be compensated by visual reward cues, suggesting that visual perception might be preserved. Ventral pulvinar (vPULV, purple shading) inactivation leads to change detection deficits resembling visual neglect. Summarized from local inactivation studies in monkeys (Wardak et al., 2002; Wilke et al., 2010; Hwang et al., 2012; Wilke et al., 2012; Wilke et al., 2013; Hwang et al., 2014; Christopoulos et al., 2015; Katz et al., 2016; Zhou et al., 2016).
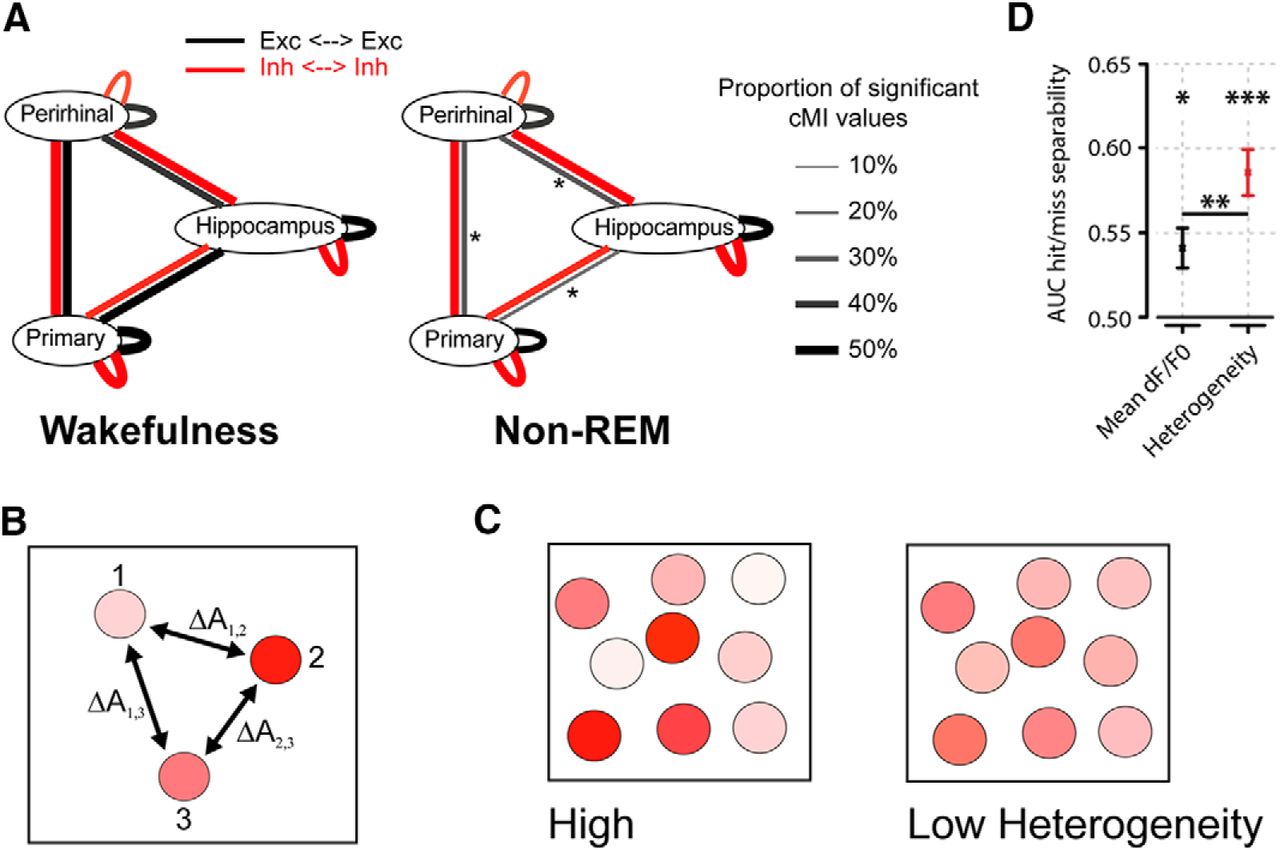
A, Summary of main findings on spike-based functional connectivity in rats (Olcese et al., 2016). Coupling was measured as pairwise cMI between single neurons. During wakefulness, cMI between neurons located in the same or different areas is largely balanced (left) for both excitatory and inhibitory neurons (black and red lines, respectively). In NREM sleep, interareal (but not intra-area) coupling between excitatory neurons is significantly reduced. This did not apply to intra-area cMI and interareal cMI (between interneurons). Line thickness indicates the proportion of neuronal pairs for which cMI was significantly >0. Asterisks indicate which connections showed a significant change between wakefulness and NREM sleep (the only significant differences found pertained to interareal coupling between excitatory neurons). Thus, during NREM sleep, neural computations may continue in local “islands of activity,” whereas global integration capabilities are reduced. B, Calculation of heterogeneity across a neuronal population (compare Montijn et al., 2015). A measure of a neuronal activity change (A, e.g., the relative fluorescence response of a neuron in 2-photon imaging, dF/F0) is computed across all neurons. Next, the responses are z-scored per neuron across all trials and all trial types (e.g., in a given session, visual stimuli are presented at 6 different contrasts; each contrast is presented 20 times; 120 trials in total). Per trial, the absolute difference in z-scored activity is then calculated across all pairs of neurons (e.g., ΔA1,2 is the difference between the responses of neurons 1 and 2). The population heterogeneity in a given trial is the mean of activity differences across all pairs. C, Examples of high (left) versus low heterogeneity (right) in a neuronal population, where response strength is indicated by color saturation. D, In a visual stimulus detection task performed by mice that were subjected to 2-photon imaging of V1 neuronal populations, heterogeneity was better capable of separating hit (detection) and miss (nondetection) trials than the mean fluorescence response (area under the curve resulting from receiver-operating characteristic analysis). Both measures predicted response behavior above chance: mean response, *p < 0.05; heterogeneity, ***p < 0.001; area under the curve = 0.5, chance level. Behavior was predicted better by heterogeneity than mean response (**p < 0.01). Values are mean ± SEM across animals. Data from Montijn et al. (2015).